近来有关人类起源的故事重新引起舆论界的注意。在2011年美国公共广播(National Public Radio)和《今日基督教》杂志(Christianity Today)都发表头条新闻, 报道某个基督徒学者不但宣称人从一个猿猴般的祖先进化而来, 而且强调科学已经驳倒了传统基督教所认同的第一对人类祖先亚当夏娃的存在?。 (注1) 他们显然已被新达尔文主义所折服, 认为我们的来源再不需要其他理论来解释。舆论界显然也相当重视这个项目, 因为这些发言人是基督徒, 想必不会对宗教持有偏见, 而且起码他们中间也有可信任的科学家。
当我初次看到这些报道时, 我很诧异这些人怎么不加批判地接受人类进化的科学论据。这是一个错误。科学并不是没有错误, 因此我们需要仔细地评估科学论据。当它涉及一个高度敏感像人类进化的问题时, 更应如此。
大部分支持人与猿猴有共同祖先的科学论据都是基于结构上的相似性, 和基因序列的相似。但是从我个人的研究结果来看, 我知道我们不能倚赖两种复杂结构之间的相似性, 来指引它们之间进化的路径。
相似本身没有提到什么机制, 能够解释二者之间是否有关系, 尤其是当二者之间的进化必须经过大量的遗传改变。事实上, 当鼓吹进化论的生物学家提出物种进化理论时, 他们忽视了实际完成物种跃迁过程所需要的遗传变量, 以及所需要的时间。实在令人惊讶! 正如我会在下文解释, 这些障碍是人类进化的一个重要的因素, 我会指出, 我们不能借着任何无引导式的过程从一种人猿祖先进化而来。
共同祖先的证据
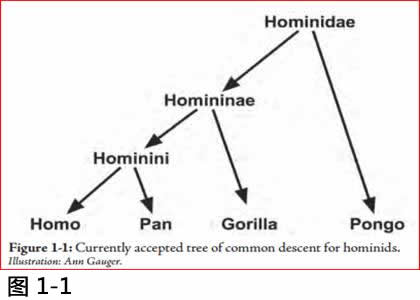
虽然达尔文时代还未发现过渡性的生物化石, 他本人已提倡人类是从一种人猿祖先缓慢进化而来的。此后, 古生物学家和人类学家已发掘到似乎是人猿之间的过渡化石。根据这些化石和活着的物种之间基因序列的比较, 他们提出图1.1中人猿有共同祖先的进化树(人与猿都同属于人科[hominids])
图1.1: 目前公认的人科共同祖先的进化树
Illustration: Ann Gauger
这树的末枝代表活着生物的属(genera, 有相似特征的物种species), 树的分叉处代表属与属之间的假想共同袓先。这树最后的分叉被称为Hominin (hominins), 包括人类和黑猩猩(Pan, Chimpanzees), 和其间的一切过渡物种。(注2)
支持这个进化树的证据是什么? 正如我上文所言, 它取决于两种数据: 巨猿, 人科化石和人类之间结构上的异同(anatomical similarities and differences), 和活着物种之间的基因序列比较(comparative analysis of DNA sequences) 。同时也建基在一个很重要却未经证实的假设: 一切可被发现的相似性能都是来自一个共同的袓先。我在这一章中就是向这个假设作出挑战。
支持这个进化树的化石证据实际上是十分简略薄弱(注3),古老的人科化石是罕有的, 所发掘到的通常是在不同区域和不同地层中找到的碎骨或脱节的骨架。这些数据可以分为两个基本类别: 仿猿或仿人的化石。它们之间的不连续性是人所共知的。著名的进化论生物学家迈尔(Ernst Mayr)在他的《什么使生物学独特? 》一书中承认它们之间有鸿沟:
最早出现的Homo化石,Homo rudolfensis和Homo erectus,与Australopithecus之间有一道很大的不能跨越的鸿沟。我们如何解释这看似跳跃的现象?既然没有任何化石作为遗失的环节,我们必须退求其次,那经时间考验的历史科学方法,就是重建一个历史的叙述。(注4)
这个历史故事就是众所周知, 在坊间杂志如《国家地理》National Geographics刊登的人类进化过程。
基因序列比较的数据也是同样的莫名其妙, 基因序列乃是由千亿万个核苷酸(nucleotides)连成的链。这些基因序列链的比较是一件棘手的事, 因为这些比较, 有可能包括个别核苷酸的兑换, 插入或删除, 基因的重复和重组, 使整过程复杂化。 (注5) 生物之间的相关程度, 有赖于科学家怎样比较这些基因, 和什么数据被包括在内或排除在外。 (注6) 但姑勿论我们与黑猩猩是多么相似, 问题的症结是: 相似性证明了什么?
对大多数的生物学家来说, 人类与黑猩猩的相似性被假设为我们源自共同袓先的证据。这个假设就是所有进化理论的基础。但请注意结构和序列的相似性本身, 并不能确认共同的血统。"野马"(Mustang)和"金牛座"(Taurus)汽车也具有很强的相似性, 因此你可能会说, 它们源自一个共同的祖先: "福特",但这些车之间的相似性是基于共同的设计, 而不是共同祖先的结果。
要是对有关共同的祖先任何故事进行验证(包括人类共同的祖先所提出的故事), 科学家必须证明两件事: 第一, 在祖先形式与新的形式之间, 必须有逐步渐进自然选择途径的存在: 无论新的形式是一个新的基因, 一个新的蛋白质, 或者一个新的物种; 第二, 如果该路径是由无引导的, 无目的性的新达尔文主义机制所操纵, 那么就必须有足够的时间和概率资源, 可被用来完成该旅程。新达尔文主义中的基因突变, 重组, 遗传漂流, 和自然选择的机制, 必须在可利用的时间内, 足以产生所提出的生物创新。要证明新达尔文进化是否已经发生过的事实, 达成这样两个条件是绝对必要的: 一个逐步自然选择的路径, 和有足够的时间和概率资源完成旅程。
然而, 在任何有关进化过渡的重要理论中, 这两件事还没有被证明过。在下文我将会表明, 这两件事情在人类进化过程中都没有得到证实, 而且可能永远不会被证实。
一个实验测试
人类真的由新达尔文主义的机制进化而来的吗? 我们无法直接查验过往, 我们只能在实验室中验证更简单的转换的可能性。
看来相像的蛋白质都被认为有一个相同的进化祖先。要是这些蛋白质有不同的功能, 那么它们当然地被认为是基于某种新达尔文主义的过程, 导致了它们的重复和分歧。这就是共同祖先的故事缩影。但是蛋白质不像人类与黑猩猩, 它们可以很容易地在实验室中被操纵, 成功地进行功能变化的考验。我们实际上可以肯定一个蛋白要切换到新的功能, 需要多少个突变, 从而确定哪些符合新进化论规则的技术创新是可能达到的。如果新达尔文的故事在这里失败了, 它就不能在任何地方成功。
图1.2: 大肠埃希氏菌(E. coli)的两个结构相似的蛋白质, Kbl 和 BioF.
Illustration: Ann Gauger and Douglas Axe.
我的同事阿克斯和我一同研究的项目是上述两个看起来很相似, 但却具有明显不同的功能的细菌蛋白质。因为它们有相似的结构, 被认为是源自百万年前一个共同祖先进化而来的兄弟。这就是图1.2中的蛋白质, Kbl 和 BioF。 Kbl 和 BioF不是彼此的直系亲属; 然而如果新达尔文主义是真实的, 从类似Kbl到类似BioF的东西的功能转变必须是可能的。此类在相似的蛋白质族群之间的功能转变比比皆是, 所以应该是比较容易实现的。
但是我们的研究结果是最少必须有七个突变, 才能使一个酵素(注: 或酶,蛋白质的一种) 经过功能转变成为另一个酵素: 这是太多的突变, 不可能在一个无引导式的新达尔文主义过程中发生。(注7)
细菌在研究进化论的实验中是个遗传工厂, 尤其是在只需要一两个个突变的个案中更为显著, 因为它们能迅速地适应环境。三个一同产生的协调中性突变(neutral mutations对生物无害无益)在细菌中也是罕见的。但是在我们的个案却需要七个或更多的突变, 才能使它们的功能兑换。在细菌族群中七个协调中性突变同时发生所需的时间, 是在1027年左右。在某种角度来看, 宇宙大约只有1010年的年份。(注8) 这个事情不可能发生。
但是这个过程正是新达尔文主义用作解释构造相像的蛋白质。如何借着突变和自然选择, 分歧演变成为有不同功能的品种, 要是演变成为有不同功能的现象, 不可以被已知的新达尔文主义过程来解释的话, 那么我们就需要其他答案。 (注9)
或许要解答你可能有的疑问: 我们的结果与一般已发表的蛋白质演变出不同功能的研究相仿。在实验室中要使蛋白质演变出真正不同的功能, 通常需要8个或更多的突变。这是超乎新达尔文主义过程的范围。
进化到人类
我在上文所说的研究指出结构的相像, 不足以建立两个有不同功能的蛋白质之间的进化途径。事实上, 新达尔文主义过程不足以产生足够的创新, 来引致那么多所需要的突变。现在我们要考虑的, 是究竟这种分析能否应用在人猿祖先进化为人所需要的演变。
首先让我们考虑我们与巨猿的分别。什么是我们鲜明的特色? 当然我们有重要结构上的分别: 我们的直立行走, 较长下肢和较短上肢, 肌能的分别, 显著较大的脑袋和头颅 (比巨猿大三倍), 和我们手、唇和舌上精致的肌肉。更重要的是我们的智能和经验, 使我们成为独一无二的人类。抽象的思考, 艺术和语文: 这些都是使我们从低等动物分别出来的特色, 不仅是程度的差异, 而且是基本上的不同。
要产生这些创新需要多少个突变? 我们缺乏衡量智能演变的数据, 所以让我只考虑我们与黑猩猩结构上的分别。
黑猩猩适合在树上生活, 我们适合在地上生活: 走路和跑步。从树上演变为完全在地上生活需要很多结构上的改变。有效的走路和跑步要有新的向骨盆倾斜的脊骨, 腿向臀部倾斜的角度可使我们的脚在走动时不致东倒西歪。我们需要为直立行走而设计的身形: 膝盖、脚、脚趾, 和平稳地坐落在脊骨上的头颅。 (我们头颅上的圆顶, 是因为要适应我们更大的大脑而向后倾斜, 但仍然保持平衡。)
若要允许人类有可以看到前面的目光, 同时仍然能够看到自己的脚在哪里, 我们的下巴和附属的肌肉必须被转移, 我们的脸变平, 脸部背后的鼻窦和眼窝, 都要位于不同的地点。
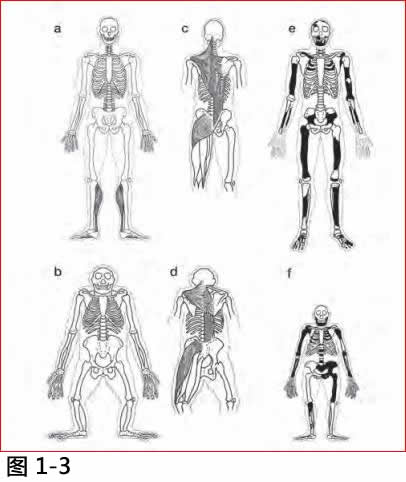
图1.3: 人科解剖结构的比较. 图示人类的(a)骨格和(c)主要的肌肉与黑猩猩的(b)骨格和(d)主要的肌肉. 以相同的比例画出的是直立人(H. erectus)(e)和南方古猿(A. afarensis)(f), 其中纯黑色的部分是已经找到的化石骨骼, 白色的部分是假想的.
Illustration: Adapted by Jonathan Jones with permission from Macmillan Publishers Ltd: Nature, Dennis M。 Bramble and Daniel E。 Lieberman, "Endurance running and the evolution of Homo," 432 (2004): 345?352, Figure 3, copyright 2004。
图1.3显示很多不同之处. (注10) 人类(a, c)和黑猩猩(b, d )有显著不同的肩膀、肋骨腔、脊骨、骨盆、臀部、腿部、手臂、手和脚。每个结构都是适合于二者不同的生活模式。
现在让我们来研究一下根据化石而来的进化故事。图中人类和黑猩猩旁边的就是从部分骨骼遗骸重建的两个人猿化石: (e) 160万年前的直立人(Homo erectus) "Turkana boy"样本, 和 (f) 320万年前的南方古猿(Australopithecus afarensis) "Lucy"样本。 (注11) 在该图中虽然"Lucy"的腿和骨盆暗示她曾花时间直立行走, 但在大多数方面她像黑猩猩, 科学家辩论她到底拥有多少陆地上的生活方式, 然而她的骨架的某些元素会减低她步行的效率。
对照之下"Turkana boy"与现代人类的结构更为相似。他所属的人科直立人(H. erectus)在若200万年前首次在化石中出现。那时已经有适合包括长途奔跑的直立体形。他与智慧人(Homo sapiens, 现代人)唯一显著的分别就是他的头颅。虽然远远大过南方古猿, 却小于现代人(但是仍然在现代人遗传变异品种范围之内)。
若我们与黑猩猩有共同袓先的故事是准确的, 要进化成为现代人应该经过像是从南方古猿到直立人的过渡程序。这里就是中断的地方。直立人就是第一个差不多以现代人的结构在化石中出现, 而且具有诸多早期的人科化石所缺乏的特征。南方古猿到直立人之间根本就没有连接这鸿沟的过渡化石。正如威斯康星州立大学古生物学家贺克斯(John Hawks, University of Wisconsin at Madison)所言:
没有南方古猿与直立人两个物种之间的明显过渡化石……我们对此的解释是, 变化是突发性和相互关联的, 反映出的是一个瓶颈: 是由一小群从南方古猿物种袓先隔离的群体所形成的。在这个人口不多,漂移和选择的组合导致对偶基因频率一个根本的转变,从根本上转变了自然选择的适应机制; 换言之, 这是一个遗传的革命。" (注12) [我加入着重之处,也为了清楚的原故删除了内部的注引。]
时间有限,要做的事情却很多
从我的论据立场而言, 我不打算争辩直立人是不是第一个人, 或者是否直接是我们血统的一部分。相反, 我希望把重点, 放在从南方古猿过渡成为直立人时, 在结构上必需要有的改变。因为无论其间有没有其他过渡化石, 这些结构的改变一定要发生。
若如贺克斯所言,有"一个根本的转变", 要借着纯粹的新达尔文主义过程发生的话, 一些突变、漂移, 和选择的组合, 一定要能够成就这事。但是从南方古猿过渡成为直立人时, "转变了自然选择的适应机制"需要转变多种解剖结构, 这过程必须有多项特异性突变。
两个问题就出来了: (1) 有多少突变会使一个南方古猿物种改变成直立人?(2) 若是从南方古猿改变成直立人所用的时间不过是150万年, 新达尔文主义的机制能否达成这些必须有的变化?
究竟这个过程需要多少个突变? 布兰布尔(Bramble)和利伯曼(Lieberman)计算在直立人或现代人首次出现的人体特征有16个。(注13) 这些功能是必要的, 用以稳定头部, 允许躯干与头部和臀部的互对旋转, 稳定躯干, 以及吸收跑步时的震荡和传递能量。许多这些的变化, 都必须一起发生, 才能有任何好处。
是否有足够的时间, 足以借着新达尔文主义过程, 来获得16个结构学上的变化? 所有这些新功能可能都需要多次突变才可达成。在实验室中要使一个细菌获得一项功能所需要的中性突变, 所能产生的极限是六个。 (按: 若超过则会有害于细菌的健康发展)。在灵长类动物(primates包括猿猴和人类)有更严格的限止。若在一个族群中要获得一个有利于进化的突变, 人类比细菌需要极长的时间, 因为据估计人比细菌的有效族群小(effective population size按: 容许基因进化的群体, 人:一万; 细菌: 十亿), 人类代际时间是十几、二十年, 细菌却能在一年内衍生千万个世代。
要是你不相信我的话, 在2007年, 达雷特(Durrett)和施密特(Schmidt)在《遗传学》(Genetics)杂志上发表一个估计, 若在灵长类动物血统中一个在核苷酸的与蛋白质衔接点(nucleotide-binding site, 按: 用作基因启动之据点)所发生的一个突变,(注14), 可被进化过程采用(注15), 需要经过长达六百万年的时间。若第一个突变是中性的话(按: 不能助长进化), 这两位作者估计还需要二亿一千六百万年, 才可以在这衔接点上达成有两个突变的机会。( 注16)
面对现实
但是普遍从人类和黑猩猩共同祖先进化成为现代人所有时间的标准估计, 是只有六百万年。二亿一千六百万年前是哺乳动物首次出现的三叠纪(Triassic)。上述所言所需要的16个结构上的改变, 实在无法在这段时间中采用一两个突变来达成, 最多只能做成一个新的蛋白质衔接功能来调节一两个基因而已。达雷特和施密特承认这个问题的存在, 但他们认为可以有二万个调节基因各自突变, 可以帮助进化, 解决这个难题。
但是这个假设是不合理的。有二万个调节基因各自突变不能减低问题的严重性。在直立人身上很多结构上的改变需要同时发生才能起作用。个别的突变是无用或是有害的。就算一两个随机而来的突变引致一个结构上改变, 它也不大可能被保留。要靠着一个无引导性的过程, 在六百万年之内, 更可能是在一百五十万年之内(按: 化石数据), 达成所有16个结构上的改变是不可能的。
用一个生活上的例子来说明: 假设让你蹒跚学步的孩子在电脑键盘上乱搅计算机的操作系统, 任凭她将1改变为0, 删除一连串的111……或0000……, 或是将它们重新排列。她能够做出一个能改进计算机的操作程式的机会有多大? 除非你有先见之明, 编写了一个执行程序, 可以把一切减少操作系统效率, 或使它崩溃的变更删除。否则你的孩子一定会把计算机的操作系统弄糟! 即使有了消除导致崩溃的变化和奖励效率的执行程序, 孩子的随意变化, 极不可能创建一个新的运作程序。这是因为执行程序没有先见之明, 无法预料某些变化如果被保存下来, 最终可能导致一个有价值新的运作子程序。
蹒跚学步的孩子就像是突变, 自然选择像是执行程序, 自然选择可以消除导致崩溃的变化并奖励效率, 但是它在创新方面是非常糟糕的。它没有先见, 不能预料那些变化可以引致创新而将它保存下来。它缺乏意愿。其实自然选择往往在当时所得的小利益之下, 放弃大量的遗传信息。(注17)
请记住细菌虽然繁殖快捷并有庞大的族群, 但需要发生六个上突变的创新是不可能的。对于大型哺乳类动物和我们一样而言, 为新达尔文主义创新的机会, 是非常非常黯淡的。
若要进化达成行走和奔跑必须的结构需要多少个突变? 如果它们可以通过随机突变而发生的话, 答案是几十个甚至几百个或者数千个。从人类和黑猩猩共同祖先要进化成为现代人, 在所有的化石数据间距六百万年的范围之内, 假设有效族群是一万, 与黑猩猩相似的共同祖先繁殖换代需时五至十年, 和每个繁殖世代中的每个核苷酸根基突变速率是一亿分之一 (10-8), 那么我们只能期望一个在蛋白质衔接位置的基因突变可能发生。然而新达尔文主义声言: 所有的16个构造特征的演变, 是偶然地在同一时间内发生, 尤其是在每一个特征的演变, 都需要多个突变。这种说法没法令人心服。若我们考虑到这些数字, 我们根本上极不可能, 或是绝对不可能, 从与黑猩猩相似的共同祖先, 借着一个渐进的, 非引导式的过程中演变而来。
人类例外
上述说法完全是基于构造上的变化, 可以达到完全直立的两足动物, 和有效的长途旅行的姿态。但是在离开这个话题之前, 我要指出人与猿之间还有很多区别。在精密机械的层面, 我们有很多功能与结构特征是猿猴所没有的——我们的手, 脸和舌头等, 具有更多有精细控制的肌肉。要是没有它们, 我们便失去作为艺术家或工匠的身手灵活性, 我们便缺乏交谈的能力, 我们也不可能用我们的面部表情, 来表达情感的细微差别。
更重要的是我们具有认知和交际能力, 我们是远超于一个有精细动作控制可以直立行走的猿猴, 我们有抽象的思维, 自我意识的反思, 和彼此相交的能力, 使我们属于一个完全不一样的类别。这些属性有更复杂的幅度, 远超于任何动物所能做到的。例如, 语言既需要身体结构上的特征 (我们的咽喉, 及在我们的大脑语言中心的构造), 还需要我们大脑中奇妙的先天而来的文法规则。三岁的小孩子都有此能力, 猿猴却是没有。真正的语言需要抽象的思维, 字句是事物和意念的代表。我们借着字句的复杂排列来彼此相交。我们有崭新的的思维, 还将这些思想传递给他人。我们会自我反省。我们会慎终思远, 吟诗作对, 研究所在的世界, 也憧憬理想中的天地。语言既反映又充实了我们的抽象思维和创造能力。
这些大量增加精细的运动灵巧性能, 以及语言、艺术和抽象思维, 在人类的快速飞跃式发展, 究竟是从何而来? 我们人类独有的特性不单是一个创新, 它构成了一个质的飞跃, 不可能借着非引导式的过程达成。(注18) 我们不是一群改装齐备的猿猴。
解释我们的来源需要处理数据的新方式。无论我们与黑猩猩有多少的相似, 从黑猩猩般的祖先演变成为人的过程, 没有严格遵从新达尔文主义的路径: 随机突变,自然选择和遗传漂流的机制, 是不足以在规定时间内, 完成所需要的变化, 所以我们需要探讨其他的解释——难道我们是某种宇宙发展的必然产品吗? 是不断枝分宇宙系列中的幸运结果吗?或者我们是一个 (些) 未知的代理人以智慧设计所实施的方案?
在评估如何解释像我们自己的生命的来源时, 我们需要选择一个能够胜任的理论。我个人相信, 没有被引导的愚蠢进程是不可能做成这工作, 这不仅是因为新达尔文主义机制是完全不足够的, 还因为我们是有智能和创作力的生命。这些素质使我们成为人类, 并与我们的能力, 同情心和我们对真善美的向往, 这些特征指引我们, 去寻找足以解释我们起源的那种原因。
图1.4:人类的创造力, 艺术, 和智能的运用是特殊的.
Illustration: Annbale Caracci, "Studio di nudo maschile," public domain, reprinted from Wikimedia Commons.
注释:
1. See, for example, Barbara Bradley Hagerty, "Evangelicals Question the Existence of Adam and Eve," National Public Radio, August 9, 2011, accessed March 6, 2012, and Richard N. Ostling,"The Search for the Historical Adam," Christianity Today, June 2011, accessed March 6, 2012.
2. The tree was recently redrawn-and the terminology changed-to accommodate sequence data that (mostly) places us in our own group with chimps. Previously the same group was called the hominids, but that term now covers all great apes and us. Some articles still use the older terminology. See http://news.nationalgeographic.com/news/2001/12/1204_hominin_id.html.
3. For more details on the subject, see chapter 3 on "Human Origins and the Fossil Record" by Casey Luskin later in this volume.
4. Ernst Mayr, What Makes Biology Unique? (New York: Cambridge University Press, 2004), 198.
5. For a discussion of one kind of rearrangement that is often used as evidence for common descent, see chapter 4 by Casey Luskin on "Francis Collins, Junk DNA, and Chromosomal Fusion."
6. T. C. Wood, "The chimpanzee genome and the problem of biological similarity," Occas Papers of the BSG 7 (2006): 1-18; G. Glazko, et. al., "Eighty percent of proteins are different between humans and chimpanzees," Gene 346 (2005): 215-219; J. Cohen, "Relative differences: The myth of 1%," Science 316 (2007): 1836.
7. A. K. Gauger and D. D. Axe, " The evolutionary accessibility of new enzyme functions: A case study from the biotin pathway," BIO-Complexity 2, no. 1 (2011): 1-17.
8. Ibid.
9. Douglas Axe amplifies this story to underscore the insufficiency of the neo-Darwinian engine to drive evolutionary change in the next chapter.
10. D. M. Bramble and D. E. Lieberman, "Endurance running and the evolution of Homo," Nature 432 (2004): 345-352.
11. "Lucy" is 40% complete as a skeleton, with only a thigh bone and partial pelvis to reconstruct her lower limbs, while "Turkana boy" is missing only the hands and feet.
12. J. Hawks et al., "Population bottlenecks and Pleistocene human evolution," Mol Biol Evol 17 (2000): 2-22.
13. Bramble and Lieberman, "Endurance running." For a list of hundreds of phenotypic traits in humans that differ from the great apes, see A. Varki and T. K. Altheide, "Comparing the human and chimpanzee genomes: Searching for needles in a haystack," Genome Research 15 (2005): 1746-1758.
14. A nucleotide-binding site is a piece of DNA eight nucleotides long. Durrett and Schmidt (see below) calculated how long it would take for a single mutation to generate a seven out of eight match for an eight nucleotide binding site (with six out of eight nucleotides already correct) in a stretch of DNA onethousand nucleotides long. Creation of such a binding site might affect the behavior of genes in the region, thus affecting the phenotype of the organism.
15. R. Durrett and D. Schmidt, "Waiting for regulatory sequences to appear," Annals of Applied Probability 17 (2007): 1-32. The relevant information appears on p. 19, where the time to fixation is factored in.
16. R. Durrett and D. Schmidt, "Waiting for two mutations: With applications to regulatory sequence evolution and the limits of Darwinian evolution," Genetics 180 (2008): 1501-1509.
17. A. K. Gauger et al., "Reductive evolution can prevent populations from taking simple adaptive paths to high fitness," BIO-Complexity 1, no. 2 (2010): 1-9, doi:10.5048/BIO-C.
18. For a review pointing out unsolved conundrums concerning our uniqueness, see a recent review by A. Varki et al., "Explaining human uniqueness: genome interactions with environment, behavior and culture," Nature Reviews Genetics 9 (2008): 749-763.