生物学家理查德. 道金斯是一个直言不讳的无神论者, 他曾经描述生物学为"研究外表好像为了某个目的而被设计的复杂事情。"(注1) 他认为这些外表完全是骗人的。生物不需要一个发明者, 因为有一个不牵涉个人意志的强大动力, 足以完成这项生物进化的工作, 即自然选择, 就是达尔文所发现的机器, 它是盲目的, 无意识地自动显示。我们现在知道, 对于好像为了某个目的而被设计的所有生物, 包括我们在内, 这就是它们如何存在的解释。"(注2)
实验证据说服了我作其他的结论。道金斯一生花了很多时间鼓吹达尔文想像中这发动机, 当你把它放到测试场所时, 便发现它实际上并没有很好地工作。我怎么知道呢? 这是因为我和几个同事数年来一直在做这些实验。我们的研究成果,有些已经在其他刊物中作详细技术的报告。事实上描述这些细节的程度, 远远超出了非科学家的兴趣范围, 我将在这里集中从宏观的角度, 以大部分使我们感兴趣, 和能使众人了解的通俗语言来说明。
我们人类是能反省自己为何存在的有生气生物, 解答我们由来的问题, 与我们的自我形象息息相关。这个探讨是古今中外的最重要的课题之一。我们都了解这个问题的重要性, 但是要衡量科学家辩论人类的来源所用的数据时, 碍于缺乏专业知识, 大多数人都是不知所措。加上科学家执着和教条化的辩论, 更使人觉得争论者可能有超过科学范围的动机。
好消息是不是看起来那么没有希望。要是观察和理解可以确实地帮助解答我们由来的问题, 那么科学会使我们往前迈进。我不是指要倚靠某个科学权威、机构、委员会或刊物, 而是科学本身。观察和理解是科学的两个栋梁, 这两个栋梁支持或否定有争论性的辩题, 理论的兴衰最终带来科学进展, 但观察和理解会继续是科学的支柱。
从这立场作为出发点, 我要提出反对达尔文主义的一个关键性的科学数据, 是每个人都可以了解的, 并以此配合人类来由的大问题。最好的论据也是简单的。在众多技术论据中精选出辩论的核心, 看看它是否有所帮助, 这是最好的策略。我认为仔细的科学分析是决定性地否决达尔文主义。无论您是否同意我的立场, 我希望能让您有足够的资料, 可作明智的抉择。
达尔文的小机器
我和我的同事高杰最近向达尔文主义提出挑战, 就是要求达尔文的机器发明一个远比人类简单的机制, 但这机制却与人类来源有一个重要的连续。有兴趣的人士可以参考我们的详细科学报告。 (注3) 在这里您只需要明白细胞中的基因, 都携带制造蛋白质的资讯, 这些蛋白质是维持细胞正常运作的工具, 您就能了解我下面的论据。
图2.1 一个达尔文的机器不能及格的适度测试. 左边之物代表原来的蛋白质, 右边之物乃是要有新的功能的蛋白质. 我们的问题并非是左边之物如何进化为右边之物, 有精确相同的外观, 我们只是要求从左边之物的功能进化为右边之物的功能. 因为它们的外貌相似, 这个过程只应需要外观的部分转移.
Illustration: Douglas Axe.
在这些简单的假设之下, 我们只问达尔文的机器, 能否在细菌中改变一个基因, 使它能制作一个有新功能的蛋白质。因为要这个实验成功, 所以我们选择了一对应该很容易胜任的蛋白质。既然无人能够预料在实验室内如何产生一个崭新的功能, 我们彻底地搜索已知的蛋白质的数据, 在其中找出一对有相似的类别(kind), 但却有不同的特殊功能的样本, 用日常生活来比喻: 要使高尔夫球赛中的推杆(putter)改变形状成为劈起杆(pitching wedge)。这个过程创造了新的功能, 但是都是用在打高尔夫球的动作中, 不需要用上完全不相干的器材如开瓶器或卤素灯。当然若遇到一个精巧的工匠, 他会很容易地把推杆改造成劈起杆。但是达尔文的机器是简单而无知的, 它能完成这任务吗?
根据我们的实验结果, 达尔文的小机器显然不能胜任: 一定不能在地球可容许进化过程的数十亿年时期内, 大概也不能在数万亿年中完成这任务。
这个结论如何影响我们的来由? 答案是它把一个重要的限制, 安置在从相似性而来的进化推论。具体来说, 我们现在知道, 我们不能这样的推断: 只是因为A和B是非常相似的, 达尔文的引擎可以从事物A产生事物B。我们为何知道这一点? 因为我们现在已经证明了一件特别的事情: 在类似的A和B事物之间, 达尔文的引擎无法完成它们的转换过程(没有直接的途径, 下面详述)。我们也知道, 从广义上来说, 我们的挑战, 在那一方面给达尔文的发动机造成了困难。我们需要它产生一个新的功能。若我们不苛求, 只要它做到改变起初的基因却保留指定蛋白质的原本功能, 达尔文的引擎是能胜任的。但是这好像是说, 如果我们忽视了这个比赛, 比赛的失败者会被认为是一个赢家。达尔文的发动机在这种情况下的失败, 就是它的垮台, 正是因为我们要求它来证明它作为一个发明家的凭据, 因为这条件是达尔文主义最高度吹捧的凭证。
下面一个原则总结了我们的结果将有助于以后的讨论:
仅仅因为A和B是基本上相似的, 我们不能推定达尔文过程可以完成从A到B的转换。
如果首次推出的原则似乎可以有放松的时候, 这一个谦逊的肯定有资格作为一个例外。这并不是说所有的达尔文式的转变都是令人难以置信的, 就像我们所研究的是可信样本之一。这原则只是说, 只是因为A和B的相似性, 不能合理地推论A是根据达尔文式的转变成为B。
这一原则虽然简单, 但是它是对达尔文主义有巨大的影响。为了全面掌握这些理念, 你需要了解相似的概念已经成为进化论坛的核心推理。由于进化生物学家认为达尔文的引擎是已能发明了世界的一切生物, 他们的兴趣只在于发动机如何在历史中活动的资料。达尔文的兴趣, 是要将所有历代的重要生物种类, 都排列在从第一个祖先进化而来的生命树的分枝上。达尔文主义的一般发明机制本身不再吸引注意力, 他们以为早在20世纪上半叶, 当达尔文的理论被遗传学当时的理解更新时, 已经充分解决了这个问题。所有今天的进化生物学家的当前急务, 只是推定整个伟大的进化家族树的细节, 这家族关系变得日益遥远, 为此他们只需不断追求各种方法, 用于检测进化家族越来越微弱的相似性。
从相似性引申而来的理论为简单两个物种相似的程度越高,它们之间的进化渊源就越是接近。但是请注意在这个简单结论背后所摒弃的东西。生物学家既然认为达尔文的机器能够达成一切生物的崭新形态, 他们就不考虑所编列的进化家族树实在是否有道理。他们的注意力完全集中在检测编排物种的相似程度, 要说服同僚, 以为若有数据支持, 引申而来的进化渊源也是正确的。
这样的思路是没有稳固根基的。达尔文的机器达成生物创新的原动力, 乃是各生物都具有的生殖功能。要是这个生物的生殖功能不是这创新的发明家时, 那么在未能解决如何创新的问题以前, 生物之间的进化渊源应该是一个有待解答的谜。按照现有的数据, 我们发现两个相似物种的进化转变, 不可能在达尔文机器的运作范围之内, 这样, 物种相似代表进化渊源这个进化家族树的理论根基, 是摇摇欲坠的。要是这个家族树被质疑的话, 那么一切从进化树引申而来的结论也要被重新考验。
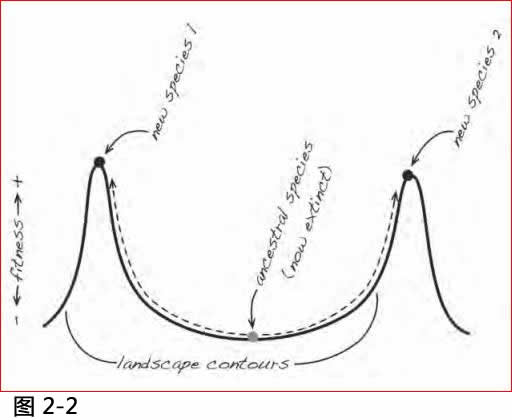
我们用一幅心理图片来说明问题的所在。达尔文进化过程常以一段冗长崎岖的景观作为比喻。这个奇特的彊域中任一个景点都代表一个可能存在的生物基因组(genome), 但是因为这些可能性多到令人吃惊, 实存的生物仅是达成这些可能性的极少数。这彊域中的地面高度是生物的适应素质(fitness), 两点间的平面距离代表生物基因组之间的差距。今天实存的百万种生物都是散布在这理念彊域中的一个任何高处。 (它们的存在代表它们基因组有充分的适应素质。)
对于在图2.2中各种实存的生物, 达尔文的引擎的倾向, 是将其移向它所能达到的最高处。根据达尔文的推论, 经过数十亿年, 这个简单地自上迁移的进化, 借着百万个分歧的路径, 使第一个原始的基因组爬上高处, 做成了今天到处可见的各种生物形式, 广布在这理念彊域中。
图2.2 达尔文对于生物多元化的解释. 这图是通过两个山峰的横截面, 代表基本上不同形式的生物. 整个景观向各个方向伸展, 拥有数百万代表所有不同种类的山峰. 这简单反复几百万次的爬山机制, 就是达尔文对于生物多元化的解释.
Illustration: Douglas Axe.
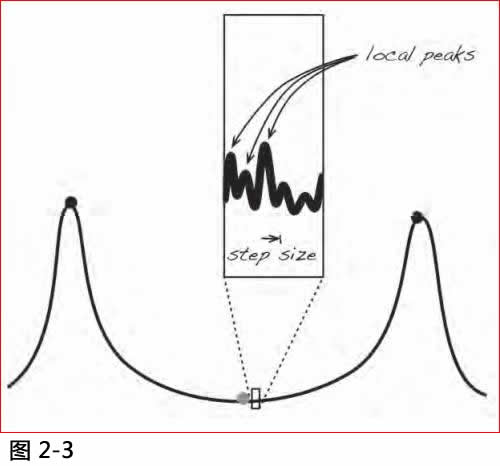
但是这个故事有一些引人怀疑之处, 在高杰博士和我做实验测试的数十年前, 一些细心的观察员已经指出这些问题(注4)。问题的症结在于距离尺度的巨大差距, 在这理念彊域中的距离度量衡, 是基于各种基因组不同的程度, 按照任何合理的计算方法, 都是非常的巨大。达尔文的发动机的进化步骤, 却只能从先前一点移动一个微小的距离。在一个步骤中, 它可以将一个基因组推到此范围的最高点, 但若要在达成任务后取得进一步的发展, 则需要更高的点坐落在达尔文发动机的活动范围之内, 偶而或可有这些机遇, 但是从第一个原始生物做成了今天到处可见的各种生物形式,要遍历所有的遥远彊域, 达尔文的发动机需要按照一个一致和有用的方法来进行, 这些机遇率实在令人惊讶!
图2.3. 攀登小步的问题. 如果发动机的动作到了可以在每个步骤中达到的最高点, 而彊域却是崎岖不平的话, 那么端点将是一个局部小峰.
Illustration: Douglas Axe.
让我们用一个较为熟悉的例子来说明, 在美国本土上的最高山峰是惠特尼峰(Mount Whitney), 高于北美洲最低洼之伯特华达盆地(Badwater Basin)只是海拔136公里。现在假设有一辆自动车, 能够远程扫描一些固定的距离内的周围地形, 然后移动到所确定的扫描内的最高点。如果扫描半径大过136公里, 这辆车就能够以一个扫描和移动的操作, 从伯特华抵达惠特尼。但是若扫描半径只有这个距离的一百万分之一会怎样呢? 现在车辆从它现有的位置"看到"四周的距离大概是一只鞋的长度, 每个移动可以达到其一半的距离。若我们考虑到十分崎岖的地面,我们不能奢望这个近视眼车前进太远, 它最多只能在被卡入岩石之前完成几个扫描和移动的操作, 也许到达离开它开始之地一半的扫描距离。它要攀登惠特尼峰根本是绝不可能的事, 所以若说任何寻求更高点的能力, 无论受到什么限制, 都能到达最高峰, 这样的想法是过分简单化的。
我们可以用与此非常相同的立论来批判达尔文主义。若达尔文的引擎可以从猿创造人类, 它就不得不在一个以单一的突变扫描为半径的严重禁区中工作。 (注5) 也就是说, 它创造人类的过程必须按部就班地以一个简单的基因突变的顺序渐进, 而每个这些突变都使其拥有者显著地比同龄者更能适应环境。若我们以这个简单的基因突变比较黒猩猩与人类基因组以百万计的分别之处, 就回到上述有关从伯特华达抵达惠特尼那个不可能完成的长途跋涉。或可假设进化的彊域比死谷(Death Valley, 按: 美国本土西部一个较平坦的盆地, 内涵伯特华达盆地)更简单和平滑, 使达尔文的发动机巡航向上在平缓的斜坡到达奇特的目的地, 但为什么会有人认为是这样的? 要证明这样神奇的事情的真实性, 一定要经过重复的实验都显示了这个种地形是规则的。
可惜的是, 我们进行的实验就是众多研究这项目的其中之一, 并且明确的共识是: 进化的彊域是不光滑和不柔和的。我们特别专注进化的发明功能, 因为这彊域的平滑度乃是这个达尔文进化机制能否成功的重要关键。从进化彊域的角度, 我们把达尔文的发动机放在自然位置(基因组的基因蛋白A), 其中具有已知的天然高峰, 而且是非常接近它的(相同的基因组以蛋白B的基因代替)。要是这个接近它的高峰有一个平缓的斜坡, 达尔文进化机制可能已经攀登一个像图2。2中较低的峰顶。但是实际上它没有这样做。虽然在科学刊物中有很多进化机制已经攀登几个高峰的报告, 但是其中并无发明新功能的例子。其实达尔文进化机制为了增加适应素质, 往往向发明新功能的相反途径迈进。(注6)
当然其他实验将继续加入这个话题。在我们的测试个案中, 达尔文的发动机不能从A移动到B的位置, 但也许一个更短距离的考验会被发现, 使达尔文引擎可以通过这个测试。对于这个问题, 也许一个在A和B之间新的蛋白质最终将被发现, 使达尔文引擎遍历中间点连接A和B的路径。重要的认知是: 这样也不会删除尺度悬殊的问题。我们现在很清楚地知道, 达尔文的发动机不能攀登相当于一个新的发明的山峰, 除非该峰恰好是非常接近它的当前位置。而且它们之间的距离小于任何一对已知有不同功能的蛋白质之间的距离。即便如此, 在非凡的情况下达尔文的发动机可以胜任, 但这将是只在这不平凡的情况下发生。要是穿越冗长的进化距离仍然依赖于一个很长的和协调特殊情况下持续发生, 这就是相当于一个不折不扣的奇迹。
事实上在我已努力地简化了这非凡的情况。在我们的实验中, 达尔文的发动机实际上比它在自然界中获得更多的有利款待, 最显著的地方是, 我们为了要在细菌细胞的运作范围做实验, 我们在从A拟转换到B的途径中设立超乎现实的安排, 事实上, 其他几个转换都必须同时进行, 而且整套运作必须恰巧都一起在同一个细胞中发生, 才能产生生物学上的意义, 从整个进化彊域的角度来看, 我们质疑达尔文的引擎与要攀升的附近山峰的距离, 实际上是比在任何现实的情形更加遥远。
从智人的高峰来看
当涉及到人类起源的理论时, 我更想补充的, 是达尔文引擎的不中用肯定更加显著, 因为它需要发明的东西变得更多。在生物界里所进行之千百万惊人的发明实例中, 没有可与发明了人类相比, 因为人类本身是发明家。如果地球上的生物进化是一场演出,萤火虫、壁虎和逆戟鲸的相继出现是壮观的话, 那么人类出场时一定会更为轰动。蟋蟀带来了更多的蟋蟀, 黑猩猩生出更多的黑猩猩。一切都非常好。人类的出现, 在另一方面, 却带来了自己的沉思产品: 音乐, 戏剧, 文学, 绘画和雕塑, 哲学和神学, 以及数学和科学技术, 体育和文化运动, 政治和战争。生物进化带来了人类最好的好处却混杂着最坏的败德, 都断然不像以前祖先中的蟋蟀鸣叫, 和黑猩猩的尖叫那么简单。
因此如果这个进化为人性化的生物有一个个自己的水平, 你想我们可以合理地归功于达尔文的小引擎吗? 我们可以说黑猩猩与人类非常相似, 以至于需要仔细解释相似的来由(很少有人会对此有异议), 但从上文可以看到, 我们虽然非常相似, 但却不能说达尔文的引擎, 已经足够越过黑猩猩与人类之间的间隙。要坚持这种立场就是忽视证据。完整的人类和黑猩猩基因组图案的比较已确定, 有20种每个都有多个基因的不同基因家族只存在于人类, 但不在黑猩猩和其他哺乳动物的遗传系统中。(注7) 当你把它与我们研究的同一家族中的基因转变比较时, 这是一个巨大差距。
事实上人类倾向接纳被重复灌输的思想, 科学家(作为人)也不例外。故事在思想框架中有其科学上地位, 但它们不能使科学如此有说服力。因此, 坚持达尔文已阐明我们人类故事的科学家们, 应该好好地思考, 支持这样的观点的证据是否有说服力。
他们有没有认真思考过从猿到人的转变究竟是怎么一回事? 他们有否想通了如何将大脑整顿为具有可以讲话或用语音的智慧? 他们知道如何在身体上配置嘴唇, 舌头和声带, 使人类可以发出语音吗? 他们有没有发现如何协调这些发明的变化和结构, 使女性可以生下大脑袋的人类后代?
如果他们用生物工程师的立场掌握了所有这些问题的答案, 他们有没有切换到遗传学家的立场, 确定了一系列单基因的突变, 作为协调整个发明人类的方法? 他们可能一开始就认为自己知道了一些问题的答案, 但是要说服像我一样的怀疑者时, 他们有否进入了灵长类动物实验室, 一直在努力工作了几十年, 悄悄地养育出可以交谈的黑猩猩, 以此来验证他们的想法呢?
如果真是这样, 他们有否做了实验, 测量最终产生的那些可以交谈的黑猩猩遗传系统中单突变的适应素质(fitness)?他们有否验证, 每个单突变增加适应素质的程度, 足以使它建立在自然的族群中(注: 即可以遗传给后代)? 假设他们已经做到了上述的一切,他们有没有从数学的角度来验证, 整个进化过渡可以在猿群几十万代内自然地发生的呢?
难以回答的问题是使人谦卑的工具, 谦卑可能是科学家赢得他们的支持者(公众) 在此话题的信任的最佳方式。事实上, 上面的清单中目前几乎没有一项是在技术上是可行的。因此我们不需要考虑任何的伦理问题。我的观点很简单: 几乎一切需要建立从人猿到人类的进化过程是否可能的实验都无法进行, 甚至在一个已经完成的科幻小说中的思想实验获得的知识,只会进一步证实, 达尔文的小引擎可以促进人类从猿??进化而来的构想, 是多么的天真无稽。
总言之, 作为一个爱好科学的人我不得不说, 人是什么这个科学课题, 是如此坚定, 如此影响深远, 如此有创意, 它应该引起每个人停下来重新考虑它的意思。我忽然想到了达尔文的答案是影响深远的, 但幸运的是, 也是彻底地错了。
注释:
1. Richard Dawkins, The Blind Watchmaker (New York: Penguin, 1986), 1.
2. Ibid., 5.
3. A. K. Gauger and D. D. Axe, "The evolutionary accessibility of new enzyme functions: a case study from the biotin pathway," BIO-Complexity 2, no. 1 (2011): 1-17, accessed March 6, 2012, doi:10.5048/BIO-C.2011.1.
4. Paul S. Moorhead and Martin M. Kaplan, editors, Mathematical Challenges to the Neo-Darwinian Interpretation of Evolution (Philadelphia: Wistar Institute Press, 1967).
5. R. Durrett and D. Schmidt, "Waiting for two mutations: with applications to regulatory sequence evolution and the limits of Darwinian evolution," Genetics 180 (2008): 1501-1509, accessed March 6, 2012, doi:10.1534/genetics.107.082610.
6. A. K. Gauger, S. Ebnet, P. F. Fahey, and R. Seelke, "Reductive evolution can prevent populations from taking simple adaptive paths to high fitness," BIOComplexity1, no. 2 (2010): 1-9, accessed March 6, 2012, doi:10.5048/BIOC. 2010.2.
7. J. P. Demuth, T. De Bie, J. E. Stajich, N. Cristianini, and M. W. Hahn, "The evolution of mammalian gene families," PLoS One 1 (2006): e85, accessed March 6, 2012, doi:10.1371/journal.pone.0000085.